前两篇讲了北疆小家鼠种群消长的4个主要外因,并指出“气候效应的实现,取决于种群自身状态”。那么,哪些内部因素对其种群消长起决定性作用呢?现场调查和饲养观察中看到,北疆小家鼠生物学特性有:小身躯、短寿命、强繁殖力、弱抗寒力、巧迁移习性、灵密度负反馈机制等等,这些都会导致其种群数量大起大落,早先各篇已有介绍。基于可定量、可先期测定,学者们考查各种动物的内源性调节因子,通常着重种群基数、年龄结构、繁殖力和性比等方面。我们对北疆小家鼠这4方面各作不同组合、不同时段的数理分析,发现前3项的作用最强、并具有该种群的特点。
开春基数(M4)——当年种群增殖的数量基础
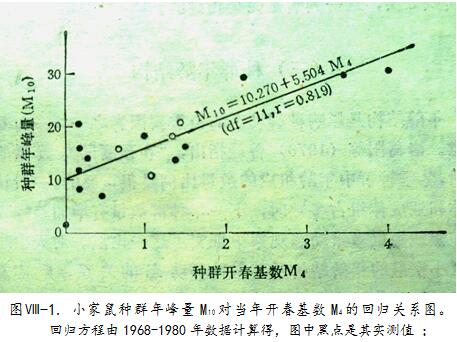
北疆全冬酷寒,田野小家鼠数量激剧下降,下降率通常在80%以上,甚至可降99.4%,13个冬季平均降88.8%。3月中下旬开春后,在房舍和野外沟渠、茬地等处过冬的存活鼠逐步聚到返青的冬作物地,田野鼠密度才回升。每年4月中旬田间捕获率反映种群的越冬存活度和数量,它们几乎全是成年鼠,是本年度的繁殖主力和增长起点,故称为“开春基数(M
4)”。这些鼠,自身既能生殖许多窝(注:人工饲养一整年的总窝数平均达9.4窝),其子孙数代亦可参与当年的繁殖,可说起着“以一当百”作用。
所以,M
4代表着当年种群增殖的数量基础,会直接制约该年种群数量水平。以1968-1980共13年的M
4,对同年“年峰量M
10”计算相关系数,r=0.819;而M
4对“年均量M
”(指4、6、8、10月共4期的捕获率平均值,代表全年数量水平),则r=0.851。两个r都>r
0.01,为极显著正相关。
由图 -1可见,M
4>0.5时,线性关系极明显,足见开春基数高是种群数量上升的重要条件。
入冬期壮龄比(L11)——年龄结构反映种群增长主力的盛衰
年龄结构是制约动物种群动态的极重要因素之一。一般认为冬季种群中幼体和亚成体占多数时,下年种群数量有增加趋势。但检测北疆小家鼠却见,入冬期幼年与亚成年的比重高反而对次年年峰量呈负相关!其实这是因为该高比重乃是出现于各高数量年后期,此时密度对生殖力负反馈制约十分强烈,甚至导致新生代变为低繁殖力基因型,照体能看就是“量多质差”。而低数量年反之,其后期种群里成年组比重高,生存力、繁殖力旺盛,后继种群必上升。就是说,此项“负相关”是密度负反馈效应所致,而非年龄结构的作用。
为查明年龄结构在北疆小家鼠种群中的作用,按臼齿磨损标准将夹捕鼠分为4个年龄组,再依不同组合、不同时段,与相应的种群数量指标作相关分析。结果发现,对其种群消长关系最密切的是11月中旬即入冬期壮龄比L
11。
“壮龄比”算式是:L
t= ∕(N- )= ∕( + + ),式中,L
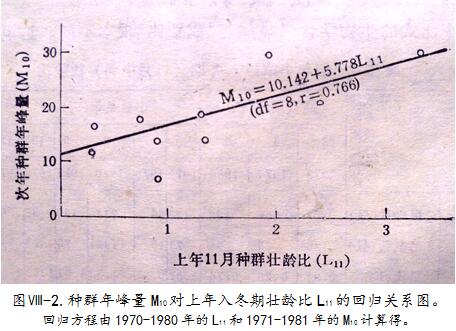
t为t月中旬壮龄比,N为该期捕获的小家鼠总数, 、 、 、 代表4个年龄组。它突出了年龄结构中 龄组的地位。就小家鼠年龄特性而言,年老、幼弱个体进严冬多难免被淘汰,能对次年种群增长起支柱作用的应是 龄组,它包括2.5月龄至1.5年龄,是生命力最旺盛的青壮个体。以1970-1980年间的L
11对各自次年的M
4、M
10计算相关系数,n=10,得r=0.665>r
0.05和r=0.766>r
0.01,为显著和极显著正相关。图 -2描述L
11与次年M
10的线性回归关系,各实测点对回归线多很贴近。
高峰期繁殖指数(f10)——秋末生殖力状况传递密度负反馈抑制烈度信息
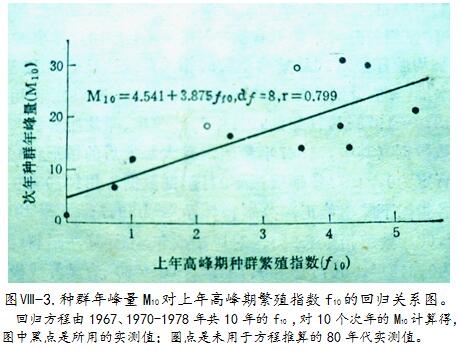
以1970-1978年加上大暴发年(1967),共10年的10月中旬捕获雌性成体(体重≥12克)之平均胎仔数、怀孕率和繁殖指数3套数据,分别对应翌年M
10计算相关系数(df=8),r值为0.711>r
0.05、0.818>r
0.01、0.799>r
0.01,为显著和极显著正相关。繁殖指数代表种群繁殖强度,其算式为:f
t=平均胎仔数 怀孕率=[怀有的胎仔总数/孕鼠数] [孕鼠数/雌成鼠总数]=怀有的胎仔总数/雌成鼠总数。(式中,怀孕率去百分号,鼠和胎仔数单位为“只”,f
t为t月份繁殖指数。)按此计算的各年10月中旬f
10称为“高峰期繁殖指数”,它对次年M
10之相关系数r值,依上述10年计算比怀孕率的r值略小,但在按不同年限计算时表现较稳定;因该指标综合了怀孕率和平均胎仔数双重信息,以它代表种群繁殖力水平更恰当。由图 -3可见,M
10 对上年f
10的回归线性关系很明显。
先前《揭秘⑷》讲述了北疆小家鼠奇妙的生育自动调节机能。秋末是种群年数量顶峰,此时种群密度对生殖力的负反馈调节效应表现最强烈,甚至新出生个体的基因型亦会改变。种群低数量年密度负反馈提升生殖力,f
10值大;随种群数量级上升,密度对生殖力抑制烈度随之增强,f
10值变小,在大暴发年后期怀孕率及f
10值降为0。由此可知,f
10实际是反映种群密度对生殖力抑制的烈度,不仅是量,而且质也变,对翌年种群增殖能力的影响尤为深刻。
性比——本种雄性偏少 与种群消长的关系主要体现在性比随密度变化
动物发育不同阶段,如岀生前后、性成熟前后,性比都会有很大变化。性别结构的变化及对数量消长的作用因物种而异。北疆小家鼠总的讲是雄少雌多,以“雄性比SR
”(即 /N= /〈 + 〉;式中, 、 分别为雌、雄鼠只数,N为总只数,比值用百分数表示)作指标,塔西河点1970-1979年田野夹捕的5656只,总性比为42.98%;并且不论分月或分年度计,各SR
值大多也低于50%。10个年度SR
平均,为43.53 2.92(%)。而依保留头骨的2945只分年龄段计,SR
分别为: 幼年组47.06%、 亚成年组56.31%、 成年组37.30%、 老年组39.35%。其中只有亚成年组是雄多于雌。再看窝仔期,该种群大多每窝5~8只仔,其SR
为47.52%。这表明,北疆小家鼠在自然出生时就是雌性偏多,但在发育为亚成体阶段雌性死亡率高,雄鼠比例可反超;到成年后,好斗的雄鼠因争雌杀伤又使其数量大减。
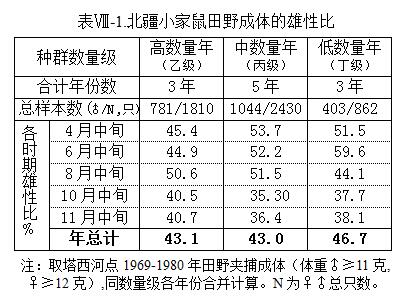
表 -1是针对成体(按体重划定, ≥11克, ≥12克,主要是 、 2个年龄组)作的统计。由该表可知,按年度总计看,低数量年的成体SR 较大(46.7%),高、中数量年则都仅约43%;一年里又有季节性变化,中、低数量级年份,上半年鼠密度尚低,雄性比也就偏大,SR 可达51.5%~59.6%,8月份开始下降,到10和11月中旬,成体雌雄比例几近2∶1;高数量年份,开春SR 就比平常年低,之后总势头也是下降。这些情形总的显示,性比随种群密度而变化,密度较低时,成体两性比例会较平衡,种群密度升高则雄性减少,但在高数量年份其季节性下降程度却又相对较小。
基于性比同种群密度的这一关联性,作反转检测:取1970-1980年(缺1971年)10年的10与11月中旬“成体雄性比SR ”数据,对其次年种群数量M4、M10计算相关系数。前3项r值未达显著,仅11月中旬SR .11对翌年M10的相关系数为r=-0.639,∣r∣>r0.05,达显著负相关水平。这是否表明性比也能反过来影响种群消长?存在两种可能的解读:
北疆小家鼠是不牢固的单婚制,生殖期内一公一母共组家庭,同生育1~2胎,到子代能独立生活时该家庭就离析,两代各个体都另行婚配形成更多新家庭;其间,一些失去生殖能力的老弱个体被遗弃,争雌行为则使雄性死亡率升高。在这种婚配制度下,春、夏季种群密度尚不高,成年两性比例接近1∶1会有利于充分婚育;入秋后,种群密度陡升,争地争食争雌的冲突加剧,此时雄性比走低,使母鼠数量相对充裕,公鼠求偶争斗缓解,减免互相伤残,应会对维持一定的繁殖水平及平稳越冬有利。上述“显著负相关”或许正是此习性的反映。
但依单婚制“一对一”特点,显然不会是“雄性比越偏低、母鼠群越偏大,越有利后继种群增长”,更不可能在中、低数量年后期成体雌雄比例降到2∶1 也对增殖有利。应当注意的是,该鼠雄性比变化到入冬期明显与前期受密度反向调节不同:数量高的年份因中期增殖数量大,亚成年期偏多的雄性此时长成,大量补入使后期雄性比下降幅度变小,SR .11值仍能保持得较好;数量低年份因增殖新个体少,雄性比持续下滑,SR .11值缩得很小。所统计的10个年度中,年均量M 秩次居前的5年,SR .11为35.1%~46.3%,平均达39.7%;秩次居后的5年,SR .11为25.0%~40.3%,平均仅34.8%。这就使得它同翌年M10表现成弱“负相关”。然而,使低数量年份的次年数量上升、高数量年份的次年数量急降,起作用最大的显然是“密度-生殖力负反馈”机制,它导致高峰期繁殖指数f10对翌年M10呈极显著正相关,分明是直接制约下年度种群消长。相形之下,SR .11效应的贡献就小得多,很可能不过是与前者效果巧合的伴生倒影。该“负相关”仅达>r0.05显著水平,而且不只成体SR .11对翌年开春基数M4、高峰期SR .10对翌年M4和M10,抑或改为亚成体SR .10及SR .11计算,各项相关系数都未达显著水平,岂不正好说明了性比对种群消长的调节作用实在不大。
三内因主导种群动向 其他因素综合调节
由上述可知,M4和L11从增长基数和繁殖主力大小两方面在数量上作用于种群年数量消长,f10则反映跨年诸个体增殖力强弱之质的差别。这3项是北疆小家鼠种群年消长的关键性内因,相辅相成地主导种群动向。在此基础上,气候、食物条件等会施以外加影响。种群消长乃是内外多因子综合作用的结果,其中“三内因”和“气候三友”既能定量,又能提前测定,可作为预测北疆小家鼠种群动态的主要指标。
但使用时须注意,各指标本身也会受别的因素干扰而失准。例如图 -1左侧各实测点很分散,就是意味着M4<0.5时种群消长更多地取决于其他因素。历年调查中观察到,M4调查之前段时期若气温特低,房舍区过冬的小家鼠会推迟外迁,若气温偏高则提早外迁,从而可影响M4的大小,使之不能确切反映开春种群总体的实际数量水平。最典型是1976年,该年4月上旬特寒,该中旬测M4仅0.16,远低于其70年代平均值(1.28),依图中所示一元回归方程计算,该年年峰量M10将仅达11.15,为“低数量级”;但该年实际越冬存活鼠并不少,到6月中旬测M6就有2.39,超过十年均值(2.23),实测该年M10=20.93,是中数量级最高年。这就是因为更多个体是4月中旬之后才由房舍里迁出之故。此例突显了综合考虑各因素的必要性,不可仅凭单个因子(M4)预测M10。再就是取样量也会影响测得指标的准确性,特别是涉及比例的,例如怀孕率、壮龄比及雄性比等,若总取样数太少,计算出的误差会很大。所以,在种群数量偏低的年份须补充捕鼠,以达到足够的样本数。