

作者:秦亚龙(江苏省中国科学院植物研究所·南京中山植物园)
监制:中国科普博览
编者按:为解码生命科学最新奥秘,科普中国前沿科技项目推出“生命新知”系列文章,从独特的视角,解读生命现象,揭示生物奥秘。让我们深入生命世界,探索无限可能。
在我国南部地区的野外或公园中,常会遇到天南星科Araceae海芋属Alocasia多年生观叶植物海芋Alocasia odora (Roxb.) K. Koch,四季常绿、株型优雅,极具观赏价值。
作为林下的草本植物,海芋的叶片为了能获取林冠透下的光斑,经历长期的进化,叶片变得越来越大,如今长宽可达一米以上。

海芋
(图片来源:作者拍摄)
在清晨或晚上高湿的特定环境下,海芋为了排出体内多余的水分,叶片边缘会“吐出”液态水滴,凝聚到叶尖后滴落,加之海芋花序形似观音,因此有了“滴水观音”的别称。在植物界,如果仔细观察,吐水现象还是很常见的。甚至,一些真菌(蘑菇)有时也会发生“吐水”现象,被称为真菌“泌滴现象”。

落叶松拟层孔菌“吐水”现象
(图片来源:作者拍摄)
海芋有毒
海芋体内含有草酸钙针晶、皂素毒苷、海芋素等有毒成分,如果不注意,海芋的汁液碰到皮肤会引起瘙痒,滴入眼睛可能会引起失明,误食海芋根茎甚至因窒息、心脏停搏而死亡。生活中偶有海芋中毒的报道,所以见到海芋时,可远观而不可亵玩焉。
在野外,有些海芋叶片上会出现规则的圆形孔洞,这是叶甲昆虫取食海芋叶片留下的杰作。当海芋遭啃食或破坏时,毒素通过叶脉系统进行释放。
虽然海芋有毒,叶甲昆虫在取食海芋叶片时,会在海芋毒素尚未传达前,快速在大叶片上啃取一圈小叶,切断毒素通过叶脉传输,然后再安心享用。神奇的是,规则的圆形是啃食相同长度下叶甲所能获取的最大的叶片面积,啃取的每一嘴都不是徒劳的。

海芋叶片上的圆孔
(图片来源:作者拍摄)
海芋花序结构和开花物候
作为天南星科植物,海芋雌雄同序的肉穗花序被保护在佛焰苞之中。佛焰苞整体分为两部分,下部分呈闭合状,卵形或短椭圆形,上部分檐部呈舟状,长圆形。
海芋花开四季,自带芬芳,花序雌雄异位,自上而下分别为附属器、可育雄花、不育雄花和雌花。附属器位于花序顶端,呈圆锥状。
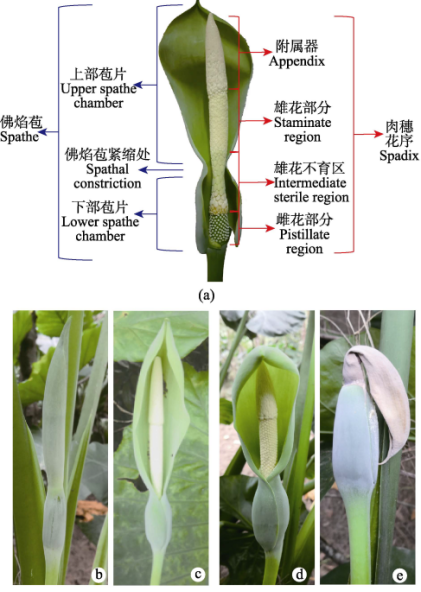
海芋花序结构和开花物候,a:花序结构;
b‒e:花序发育过程(b:花苞期;c:雌花期;d:雄花期;e:结实期)
(图片来源:参考文献)
海芋开花时,海芋的雄花负责产生花粉,不育雄花为传粉昆虫提供食物,雌花接收花粉后孕育果实。
海芋的花序还具有雌雄异熟的特征。花序中的雌花先熟、雄花后熟。花序外部的佛焰苞上部和下部苞片之间具有一个收缩处,将花序上部的可育雄花与下部的雌花进行隔离。
海芋花序具有开花生热效应。在花期,花序通过提升温度,吸引传粉昆虫并为它们提供庇护所。同时,花序升温也可以加强花香散发并促进花器官发育,这和荷花玉兰和莲开花生热现象相对一致。
在雌花期,海芋花序的花香最为浓烈,柱头黏性最强并存有少量分泌物。此时,附属器和雄花区开花生热效应显著,佛焰苞收缩处尚未完全收紧,所有访花者均可经由收缩处进入雌花区,其中就包含携带着外来花粉的传粉者帮助雌花进行授粉。
而在雄花期,花序的花香和开花生热效应逐渐减弱。此时,佛焰苞收缩处已完全紧缩,成熟的花粉和所有访花者均已无法到达雌花区,花序结果与否已成定局。

海芋浆果
(图片来源:作者拍摄)
当海芋雌花授粉完成后,随着果序的成熟,里边红色的浆果会露出。鸟类被浆果艳丽的色彩吸引来取食,后将海芋种子通过粪便排出,帮助海芋实现种子的远距离传播。
访花者与传粉者
在植物传粉期,植物通过颜色、气味和形态等花部信号,吸引昆虫、鸟类等到访。根据到访昆虫的功能不同,可分为访花者和传粉者。
访花者多是以取食花粉、花蜜、分泌物或花组织等为目的,或是在花序产卵地或偶然性停留。其中,作为花粉传递有效载体真正帮助植物完成授粉的访花者,才能被称为传粉者。
泛化传粉系统
对于那些传粉者与植物之间传粉关系不专一的,就属于泛化传粉系统。对于某种泛化传粉植物而言,多数花会产生大量的花粉,这些花粉不但能满足传粉者授粉和取食的需要,还能满足大量访花者所带来的花粉损耗,传粉效率低下,自然界多数有花植物都属于泛化传粉系统。
特化传粉系统
丁翔等最新研究指出,芋果蝇属Colocasiomyia的海芋果蝇C. alocasia和异海芋果蝇C. xenalocasiae是海芋授粉过程的高度特化传粉昆虫,它们与海芋建立起了特化传粉系统。
在雌花期,芋果蝇携带着外来花粉通过佛焰苞收缩处进入雌花区授粉,在这一过程中,以雄花不育区和雌花区柱头分泌物为食。在雄花期,芋果蝇活动在雄花可育区,进行花粉取食和交配,之后会携带着可育雄花区的花粉前往另一株海芋花序进行授粉。
在这一传粉系统中,虽然也常有露尾甲科、蜜蜂科、隐翅虫科和跗线螨科等10种非传粉昆虫访问花序,但它们并不参与授粉,而是与传粉者抢食花粉和花序分泌物,甚至是进行繁殖,从而与芋果蝇的授粉和繁殖形成竞争。

海芋常见访花昆虫及后代的活动行为:a‒c:各时期花序内芋果蝇后代活动;
a:芋果蝇虫卵(雌花期);b:芋果蝇幼虫(雄花期);
c:芋果蝇虫蛹(结实期);d:访花者将花粉传递至柱头(雌花期);
e:蜜蜂科访花者收集花粉(雄花期);f:露尾甲科访花者啃食不育花(雌花期);
g:不育花被啃食的痕迹;h:孵化的露尾甲科幼虫(雄花不育区)。
(图片来源:参考文献)
神奇的是,芋果蝇在海芋花序中的繁殖过程与海芋花序生长阶段相匹配。芋果蝇在雌花期产卵,雄花期发育成幼虫,结实期长成虫蛹。
在自然界,还有许多植物与特化传粉者建立起了特化传粉系统。
榕树—榕小蜂特化传粉系统
自然界有800多种榕树,它们与榕小蜂历经约7500万年的协同进化,逐渐形成了高度特化的互惠共生系统,成为目前已知最专一的传粉和共生系统。
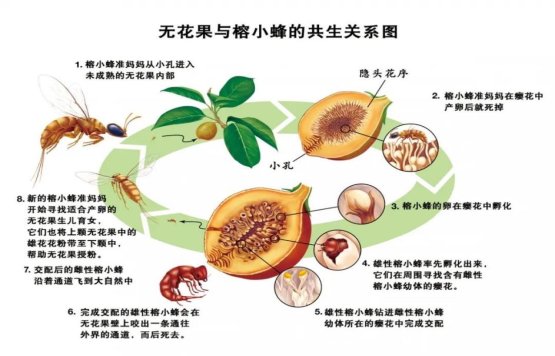
榕树独特的“隐头花序”结构,在自然界中只有榕小蜂才能帮助其传粉。
无花果与榕小蜂共生关系图
(图片来源:世界知识画报)
榕小蜂又分为传粉榕小蜂和非传粉榕小蜂,通常每种榕树只有专一的传粉榕小蜂为其传粉。虽然有些榕果内还会共存几种到几十种非传粉榕小蜂,但它们却几乎不为雌花传粉。
与海芋相似,对于雌雄同株的榕树而言,同一花序的雌花和雄花成熟期不同,处于雌花期的榕果会释放出特殊的挥发物,吸引特定的传粉榕小蜂携带雄花花粉钻进正处于雌花期的新花序内完成授粉。
凤尾丝兰—丝兰蛾特化传粉系统
凤尾丝兰原产北美,每年八九月份进入花期,大型圆锥花序白天闭合,晚上开放。凤尾丝兰的花朵没有萼片,具有6个花被片和6枚雄蕊,中间围着由三心皮联合形成的复雌蕊,雌蕊的下半部呈柱状,顶端三裂而外翻。

凤尾丝兰
(图片来源:作者拍摄)
在中国,凤尾丝兰年年开花却不见结果,这是由于缺少了北美原产地唯一的传粉昆虫——丝兰蛾。在当地,凤尾丝兰夜晚开放,散发出特有香味,吸引丝兰蛾到来。此时,到访的丝兰蛾已完成交配,雌蛾会首先飞到雄蕊顶端,采集花粉并搓成团。
之后,雌蛾带着花粉团飞抵另一朵花,停在雌蕊柱侧面,它们会用产卵器刺穿子房壁并产下1—6枚卵。产卵完成后,雌蛾还会爬到柱头顶端,将从上一朵花采集来的花粉团尽可能地通过雌蕊顶端的孔洞塞进深处,帮助凤尾丝兰完成授粉。

凤尾丝兰花朵
(图片来源:作者拍摄)
授粉完成后凤尾丝兰,子房逐渐发育膨大。丝兰蛾宝宝在其子房中孵化后,以凤尾丝兰部分鲜嫩的种子为食,最终破壁而出,钻进土中化蛹。到了来年凤尾丝兰开花时,蛹中的丝兰蛾破茧而出,寻得伴侣交配后,飞向凤尾丝兰花序授粉并产卵,开启又一次生命的轮回。
就这样,凤尾丝兰与丝兰蛾经历长期的协同进化,丝兰蛾成为凤尾丝兰唯一的传粉昆虫,凤尾丝兰为丝兰蛾的繁衍提供住所和食物,两者构建了特化传粉系统,达成了专性共生关系。
耧斗菜特化传粉系统
在生活中,还有一类颜值非常高的植物,也与传粉者建立了特化传粉系统,它们便是毛茛科Ranuncuaceae耧斗菜属Aquilegia植物。
耧斗菜的名字源自我国古时农耕播种工具——耧车,因其花瓣下部常延伸出管状的距,像极了耧车的耧腿。
耧斗菜的距内的蜜腺组织会分泌出传粉者喜欢的花蜜,但这些花蜜并不是为所有访花者准备的,只有那些帮助耧斗菜传粉的访花者,经过长期的协同演化,才可以获取鲜美的花蜜。

耧斗菜属
(图片来源:作者拍摄)
不同种的漏斗菜花距长短和形态各不相同,通过花距和花距底部的花蜜,每一种耧斗菜都会有特定的天蛾、雄蜂、蜂鸟等不同传粉者帮助其完成传粉,建立起了特化的传粉系统。
彗星兰—长喙天蛾特化传粉系统
1862年,英国博物学家达尔文收到一份来自非洲马达加斯加的兰花标本,标本的花朵有着一条长达30厘米的花距,他猜想当地一定生活着一种口器(喙)长度与之相当的天蛾能够吸食到花距末端的花蜜并为其传粉,人们因此称之为达尔文兰。因其花朵形态看似拖着长尾巴的彗星,因此得名“大彗星兰”。又因其超长的花距,其学名被定为长距彗星兰Angraecum sesquipedale Thouars。

长距彗星兰
(图片来源:作者拍摄)
直到1903年,人们才在马达加斯加发现了达尔文预言的长喙天蛾,它们拥有长达30厘米的喙。时间来到1992年,人们首次拍到了长喙天蛾为长距彗星兰传粉的过程,完美地诠释了植物与传粉者的协同进化,两者之间构建起了特化的传粉系统,共同在马达加斯加繁衍。
结语
经过长期的演化,全世界约90%的被子植物都需要昆虫、鸟类和哺乳动物等作为媒介传播花粉,它们构成了各具特色的泛化传粉系统和特化传粉系统,共同推进着动植物的进化和繁衍,持续述说着自然万物的生存智慧。
参考文献:
丁翔, 余元钧, 宋希强, 等. 具有泛化访花者的海芋特化传粉系统[J]. 生物多样性, 2024, 32 (6): 24069, pages 1-11.